





Ancient Punt, Cushitic peoples, and other Afro-Asiatic-speaking populations
The Universal Basic Genetic Model (UBGM) is a genetic model that allows one to quickly determine an individual’s ancestral composition. First introduced on Genome analysis of modern Semitic individuals, it simply consists of the main, least-admixed ancient West Eurasian, East Eurasian, South Eurasian, North African and Sub-Saharan African genome components that are known to exist.
These ancestral elements are, again, as follows:
Least-admixed West Eurasian reference samples:
IRN_Ganj_Dareh_N (Iran Neolithic component)
Levant_Natufian_EpiP (Levantine Natufian component)
AZE_Caucasus_lowlands_LN (Caucasus Hunter-Gatherer component)
TUR_Marmara_Barcin_N (Anatolian Neolithic component)
WHG (Western Hunter-Gatherer component)
Yamnaya_RUS_Samara (European Steppe component)
Least-admixed Sub-Saharan African reference samples:
CMR_Shum_Laka (ancient Pygmy component)
COG_Kindoki_230BP (ancient Niger-Congo component)
KEN_Kakapel_300BP (ancient Nilo-Saharan component)
MWI_Chencherere (ancient East African Hunter-Gatherer component)
ZAF_2000BP (ancient Khoisan component)
Least-admixed Iberomaurusian reference sample:
MAR_Taforalt (Iberomaurusian component)
Least-admixed East Eurasian reference sample:
CHN_Huatuyan_500BP (ancient East Asian component)
Least-admixed South Eurasian reference sample:
LAO_Hoabinhian (Ancient Ancestral South Indian component)
Let us now take a look at each of these ancestral elements, and recap how they relate to the Cushitic peoples and other Afro-Asiatic-speaking populations.

The Iran Neolithic component is a West Eurasian ancestral element belonging to early farmers of the Zagros Mountains in Iran. It is closely related to the Caucasus Hunter-Gatherer component (discussed below), but differs from it by containing some deeply embedded admixture associated with the Mbuti pygmies of central Africa (cf. Lazaridis et al. (2018)).
Ancient DNA analysis has revealed that the Iran Neolithic element was a core ancestral component borne by the makers of the Indus Valley Civilization (IVC). Iran Neolithic ancestry has been estimated to comprise between 45%-82% of the Indus Valley people’s ancestral makeup, with the rest mainly consisting of the Ancient Ancestral South Indian component (11%-50%) (Narasimhan et al. (2019)).
With regard to modern populations, the Iran Neolithic component attains its highest average frequencies among the Balochi, Makrani and Brahui populations of South/Central Asia:


Among Afro-Asiatic speakers, the Iran Neolithic component reaches a frequency peak among Semitic-speaking Assyrians, Sephardic Jews and Ashkenazi Jews (17.3% among Assyrians). It is generally absent among Afro-Asiatic-speaking populations in Africa.



The Levantine Natufian component is a West Eurasian ancestral element belonging to the Epipaleolithic Natufian hunter-gatherers of the Levant. It has often been linked with the ancient dispersal of the Afro-Asiatic languages and the Y-DNA haplogroup E. In Africa, Levantine Natufian ancestry has also been associated with the spread of pastoralism and the Y-DNA haplogroup T via the Middle Neolithic herders of Skhirat, Morocco (for details on this, see First genomes from the ancient Sahara detect two different groups of pastoralist inhabitants).
With regard to modern populations, the Levantine Natufian component attains its highest average frequencies among the Semitic-speaking Mahra and other Yemenis, Socotri, Saudi and Arab bedouin populations of the Middle East:


The Levantine Natufian element constitutes just over half of the predominant non-African ancestry (>70%), which Cushitic peoples bear.






















The Caucasus Hunter-Gatherer component is a West Eurasian ancestral element belonging to Neolithic period hunter-gatherers from the Caucasus Mountains. It is associated with the ancient dispersal of the Y-DNA haplogroup J.
With regard to modern populations, the Caucasus Hunter-Gatherer component attains its highest frequencies among Georgians and other communities inhabiting the Caucasus area:


Among Afro-Asiatic speakers, the Caucasus Hunter-Gatherer component reaches a frequency peak among Assyrians (56.4%).




The Anatolian Neolithic component is a West Eurasian ancestral element belonging to the first agriculturalists of Anatolia/Turkey. It is the defining ancestry of the Early European Farmers, and still comprises one of the core elements of European heritage.
With regard to modern populations, the Anatolian Neolithic component attains its highest frequencies among Sardinians and other communities native to southern Europe:

Among Afro-Asiatic speakers, the Anatolian Neolithic component reaches a frequency peak among northern Maghrebi populations, Ashkenazi Jews and Sephardic Jews (32.8% among Ashkenazi Jews of Belarus) i.e., Afro-Asiatic-speaking groups that received significant gene flow from southern Europeans.








The Western Hunter-Gatherer component is a West Eurasian ancestral element belonging to Mesolithic period foragers of Europe. Although widely distributed throughout the continent, it is a vestigial ancestry which only exists today at low frequencies.
With regard to modern populations, the Western Hunter-Gatherer component attains its highest frequencies among Basques:


Among Afro-Asiatic speakers, the Western Hunter-Gatherer component reaches a frequency peak among Ashkenazi Jews (1.6% among Ashkenazi Jews of Ukraine), due to admixture with Europeans.

The European Steppe component is a West Eurasian ancestral element belonging to nomads of the Pontic-Caspian Steppe in eastern Europe. It is associated with the ancient spread of the Indo-European languages and the Y-DNA haplogroup R1. Along with the Anatolian Neolithic component of the Early European Farmers and (to a lesser extent) the WHG component of the Mesolithic Western Hunter-Gatherers, the Steppe component forms the core ancestry borne by modern European individuals.
With regard to contemporary populations, the European Steppe component attains its highest frequencies among the Khanty, Udmurt and other communities of northern and eastern Europe:


Among Semitic-speaking populations, the European Steppe component reaches a frequency peak among Ashkenazi Jews and Sephardic Jews (21.2% among Ashkenazi Jews of Belarus), due to admixture with Europeans.
On the Universal Basic Genetic Model, which presents a rough overview of a person’s ancestral composition, Cushitic individuals show a low European Steppe ancestry of under 10%. However, more precise distal genome analysis indicates that they actually carry significant European Steppe ancestry, averaging around 20% (see Detecting the Eurasiatic genome component in Cushitic peoples). Consequently, many Cushitic individuals have been observed to bear European Steppe-associated mtDNA haplogroups, such as the I clade and haplogroup H derivatives.









The ancient Pygmy component is a Sub-Saharan African ancestral element belonging to early Pygmy-related hunter-gatherers of Shum Laka, Cameroon.
With regard to modern populations, the ancient Pygmy component attains its highest frequencies among the Baka, Bakola, Biaka, Mbuti and other African Pygmy groups:


Among Afro-Asiatic speakers, the ancient Pygmy component reaches a frequency peak among Bedouin Arabs of the Levant (0.4% among BedouinA from the Negev desert). It is generally not carried by most Afro-Asiatic-speaking individuals.

The ancient Niger-Congo component is a Sub-Saharan African ancestral element belonging to early agriculturalists of Kindoki, Congo. It represents the principal ancestry borne by Niger-Congo speakers in western Africa and their descendants, including African Americans and Afro-Caribbeans.
With regard to modern populations, the ancient Niger-Congo component attains its highest frequencies among the Esan, Mende, Yoruba and other Niger-Congo-speaking individuals in western Africa. This element is also borne at elevated percentages by the Hausa and other Chadic speakers, who today speak Afro-Asiatic languages, but appear to have originally spoken Niger-Congo languages:


Among Afro-Asiatic speakers, the ancient Niger-Congo component reaches a frequency peak among individuals in southern Morocco and other such contact zones, where Afro-Asiatic speakers have historically interacted with Niger-Congo speakers. In other parts of the Maghreb and elsewhere, most Afro-Asiatic speakers do not bear any Niger-Congo-related admixture (for further discussion on this, see Y-DNA haplogroup E1b1a (formerly E3a) appears to have originated in the Middle East).









The ancient Nilo-Saharan component is a Sub-Saharan African ancestral element belonging to early modern inhabitants of Kakapel, Kenya. It comprises most of the ancestry carried by both Nilo-Saharan and Niger-Congo speakers in eastern Africa. As such, it appears to be the genomic analogue to the linguist Roger Blench’s Niger-Saharan macrophylum, which posits that “Niger-Congo and Nilo-Saharan are genetically related and should be regarded as a single macrophylum” (cf. Blench (2008)).
Admixture runs suggest that the ancient Nilo-Saharan component is related to the ancient Niger-Congo component. However, it differs from the latter by the fact that it lacks alleles derived from archaic African specimens, which are embedded within Niger-Congo ancestry (for more on this archaic African admixture, see here).
With regard to modern populations, the ancient Nilo-Saharan component attains its highest frequencies among the Luo Nilotes and Luhya Bantus:


Among Afro-Asiatic speakers, the ancient Nilo-Saharan component reaches a frequency peak among certain Sudanese “Arab” individuals (as high as ~50% among some Kababish). However, among other Sudanese “Arabs” and Afro-Asiatic speakers generally, it occurs at low-to-moderate frequencies.











The ancient East African Hunter-Gatherer component is a Sub-Saharan African ancestral element belonging to early foragers of Chencherere, Malawi.
With regard to modern populations, the ancient East African Hunter-Gatherer component attains its highest frequencies among Hadza forager individuals:


Among Afro-Asiatic speakers, the ancient East African Hunter-Gatherer component reaches a frequency peak among Iraqw individuals in Tanzania (~20%).


The ancient Khoisan component is a Sub-Saharan African ancestral element belonging to early foragers of southern Africa.
With regard to modern populations, the ancient Khoisan component attains its highest frequencies among Khoisan forager individuals:


Among Afro-Asiatic speakers, the ancient Khoisan component reaches a frequency peak among Ashkenazi Jews (0.2% among Ashkenazi Jews of Lithuania). It is generally absent among other Afro-Asiatic-speaking populations. (For specifics, see Ancient Khoisan were dark-skinned and did not look Eurasiatic.)

The ancient Iberomaurusian component is a North African ancestral element belonging to Epipaleolithic hunter-gatherers of northern Africa, who are associated with the Iberomaurusian archaeological culture. It is closely related to the Levantine Natufian component, but differs from it by containing ~33% of Sub-Saharan African admixture (cf. Ferreira et al. (2021)).
With regard to modern populations, the Iberomaurusian component attains its highest frequencies among Berbers and other Maghrebi populations:


The Iberomaurusian ancestral component is also present at low frequencies among most other Afro-Asiatic speakers. Among Cushitic speakers, Sudanese “Arabs” and other pastoralist groups in the Horn of Africa, Nile Valley and Chad Basin, this genome element was mostly acquired via the Middle Neolithic pastoralists of Skhirat (who bore ~14% of Iberomaurusian-related admixture).




The ancient East Asian component is an East Eurasian ancestral element belonging to early inhabitants of Huatuyan, China.
With regard to modern populations, the ancient East Asian component attains its highest frequencies among Gelao and Hmong individuals:


Among Afro-Asiatic speakers, the ancient East Asian component reaches a frequency peak among Somalis and other Cushitic individuals in the Horn. It is also present at lower frequencies among most other Afro-Asiatic-speaking populations.
Along with the European Steppe component, this ancient East Asian element appears to have spread from South/Central Asia into Northeast Africa, thereby introducing Eurasiatic (i.e. European Steppe and ancient East Asian) ancestry to these areas.

The Ancient Ancestral South Indian component (AASI) is a South Eurasian ancestral element belonging to early inhabitants of the Hoabinhian cultural complex in southeast Asia.
With regard to modern populations, the Ancient Ancestral South Indian component attains its highest frequencies among the Onge and Jarawa of the Andaman Islands, as well as tribal and Dravidian-speaking groups in southern Asia. For this reason, the AASI element is also known as the Onge component (cf. Lazaridis et al. (2016), Extended Data Figure 5):


Among Afro-Asiatic speakers, the Ancient Ancestral South Indian component reaches a frequency peak among EmiratiC individuals in the United Arab Emirates (10.9%). It is generally absent among other Afro-Asiatic-speaking populations.





Genome analysis indicates that Cushitic, Ethiosemitic and North Omotic-speaking populations of the Horn region predominantly bear non-African ancestry (over 70% on average), which consists of a Levantine Natufian component and a Eurasiatic component (i.e. European Steppe and ancient East Asian elements). They also have minor Sub-Saharan African admixture (~25%) and trace Iberomaurusian admixture (under 5%).
The Universal Basic Genetic Model detects this same overall ancestral composition, albeit with less numerical precision. Below are UBGM data tables for each Horn population listed in the Global25 modern datasheet. For the exact frequencies of these various ancestral components, refer to Detecting the Eurasiatic genome component in Cushitic peoples and First genomes from the ancient Sahara detect two different groups of pastoralist inhabitants.
















Genome analysis of modern Semitic individuals
Correcting a faulty genetic model #1
Detecting the Eurasiatic genome component in Cushitic peoples
First genomes from the ancient Sahara detect two different groups of pastoralist inhabitants
Y-DNA haplogroup E1b1a (formerly E3a) appears to have originated in the Middle East
Baggara/Shuwa “Arabs”: last remaining descendants of the original Chadic speakers
Phenotype of the Canaanites, ancestors of modern Semitic peoples of the Levant and Mesopotamia
On The Dynastic Race Theory might be true, we saw how Egyptian individuals dating from the Old Kingdom period, who were analysed for ancient DNA, were found to almost entirely carry Levantine Natufian ancestry. This result appeared to confirm the Dynastic Race Theory, a hypothesis which asserts that a “New Race” from the Middle East introduced most of the Pharaonic (Dynastic) culture to the Nile Valley. Under this view, the Egyptians of the earlier Predynastic period would have been of a different, European-related ancestral stock.



I suggested that the basic tenets of the Dynastic Race theory are correct because:


Besides the above, there is some additional supporting evidence I did not touch on; namely, what seems to be Predynastic Egyptian admixture in the northern Nilotes of the Nile Valley.
On Correcting a faulty genetic model #1, I demonstrated through genome analysis how the Dinka and other northern Nilotes bear moderate non-African admixture. That non-African admixture consists of a blend of European Steppe and ancient East Asian elements, with no apparent Levantine Natufian component:

Where could this non-African admixture have come from?
It clearly was not from Cushitic speakers. Although European Steppe and ancient East Asian elements (Eurasiatic ancestry) are core components of Cushitic heritage, ancient and modern Cushitic individuals also possess substantial Levantine Natufian ancestry. Thus, any population that has received gene flow directly from a Cushitic source should carry at least some trace of Levantine Natufian admixture (e.g. the Sandawe and Maasai Nilotes, populations which today have low-to-moderate Natufian admixture due to contacts with Cushitic peoples). Because modern Egyptians, Nubians and Sudanese “Arabs” likewise bear significant Levantine Natufian ancestry, they too could not have contributed that non-African admixture.
But what about the Predynastic Egyptians? Could they have introduced Eurasiatic elements into the northern Nilotes’ gene pool? Y-DNA analysis suggests that they did.

Hassan et al. (2008) analysed various Afro-Asiatic and Nilo-Saharan-speaking populations of Sudan for Y-DNA. He reports that most of his Dinka Nilote sample belongs to the archaic African paternal haplogroup A (16 out of 26 individuals or 62%). However, a significant minority carries the V22 clade, a sublineage of the Afro-Asiatic-associated E1b1b haplogroup (4 out of 26 individuals or 15%).
In the archaeogenetic record, E1b1b-V22 has almost exclusively been observed among ancient Egyptian mummies, and often alongside the R1b haplogroup. This is telling because the R1b lineage is closely affiliated with the spread of European Steppe ancestry.
From the foregoing we may surmise that, before the arrival of the Levantine Natufian-related “New Race” from the Middle East, the Predynastic Egyptians probably carried Europe Steppe and ancient East Asian genome components. If this inference is correct, as the only people in Africa known to possess such Eurasiatic ancestry without any accompanying Levantine Natufian ancestry, the Predynastic Egyptians would be the logical choice for the population that transmitted European Steppe and ancient East Asian elements to the northern Nilotes.





The Dynastic Race Theory might be true
Correcting a faulty genetic model #1
Phenotype of the Canaanites, ancestors of modern Semitic peoples of the Levant and Mesopotamia
Genetic and phenotypic affinities of the Bronze Age Semites of Sidon, Lebanon
Detecting the Eurasiatic genome component in Cushitic peoples
As discussed on Queen Tiye, an ancient Egyptian noblewoman of Eurasiatic origin, the central tenet of the Afrocentric “Black Egypt” theory is that the ancient Egyptians were “Negroes” and African Americans/Afro-Caribbeans, the principal supporters of this fringe hypothesis, are therefore related to them. Afrocentric commentators — referred to derisively as Hoteps by non-Afrocentric African American/Afro-Caribbean individuals — assert an ancient Egyptian connection either directly via their own West African ancestors, or indirectly through Nubians and admixed Afro-Asiatic speakers.
As of 2024, the “Black Egypt” theory has been completely invalidated by ancient DNA analysis (see Guide to the Universal Basic Genetic Model). What is lesser known is that craniometric analysis has further laid to rest this far-fetched claim.
In 2006, Martha Katherine Spradley conducted a craniometric analysis of various populations from Africa. She compared the skulls of contemporary and early modern individuals from Egypt, northern Somalia, Sub-Saharan Africa, and the African Diaspora (African Americans, labeled “Enslaved” in the study).
Spradley observed that the Afro-Asiatic-speaking Egyptian and Somali samples clustered together, separately from the other populations. As the scientist notes, “Somalia and Egypt are closest to one another.”

The morphological affinities shared between Spradley (2006)’s Egyptian and Somali samples are to be expected. For one thing, they have been reported in multiple other craniometric studies (e.g. Billy (1981b); Rösing (1990); Brace (1993); Terrazas Mata and Benavente (2013)).
The real question, then, is why do these ties exist in the first place?
A cursory look at the genomic profile of Egyptian and Somali individuals would suggest that these affinities stem from common Levantine Natufian ancestry. However, if this alone were indeed the case, it would not explain why Predynastic Egyptian samples often appear to share a closer affinity with Afro-Asiatic-speaking samples from the Horn of Africa (Cushitic speakers) than with Afro-Asiatic-speaking samples from the Arabian peninsula (Semitic speakers), where Natufian ancestry peaks. For example, in Kemp (2006):

On The Dynastic Race Theory might be true, I suggest that this might be because the Predynastic Egyptians bore significant Eurasiatic ancestry (European Steppe and ancient East Asian components), like ancient and modern Cushitic speakers have been found to bear. A “New Race” carrying Levantine Natufian ancestry, arriving from the Middle East just prior to the Old Kingdom, would have introduced much of the Pharaonic/Dynastic culture. Later migrations of outsiders bearing Caucasus Hunter-Gatherer-related ancestry during the Old Kingdom-Middle Kingdom would have further shaped the Egyptian gene pool.



Guide to the Universal Basic Genetic Model
The Dynastic Race Theory might be true
Northern Somalis from Puntland (Majerteen Darod) have the most non-African ancestry in the Horn
Detecting the Eurasiatic genome component in Cushitic peoples
The contemporary Semitic-speaking populations of the Middle East are genetically and physically diverse. Most researchers have attributed this heterogeneity to differing admixture strains, which were superimposed on a shared Levantine Natufian ancestral base. A few have instead argued that this diversity exists because these groups actually have separate ethnic origins.
I wanted to find out the truth underlying these claims, so I performed a genome analysis.

On the Vahaduo Admixture JS program, for the Target samples, I used most of the Semitic-speaking individuals listed in Eurogenes’ official Global25 modern datasheet (unscaled). For the Source samples, I utilized the Global25 ancient datasheet’s least-admixed representatives for each of the main West Eurasian and Sub-Saharan African ancestral components. I also included three additional ancient reference samples in order to capture any existing Iberomaurusian, East Eurasian and South Eurasian ancestries.
Least-admixed West Eurasian reference samples:
IRN_Ganj_Dareh_N (Iran Neolithic component)
Levant_Natufian_EpiP (Levantine Natufian component)
AZE_Caucasus_lowlands_LN (Caucasus Hunter-Gatherer component)
TUR_Marmara_Barcin_N (Anatolian Neolithic component)
WHG (Western Hunter-Gatherer component)
Yamnaya_RUS_Samara (European Steppe component)
Least-admixed Sub-Saharan African reference samples:
CMR_Shum_Laka (ancient Pygmy component)
COG_Kindoki_230BP (ancient Niger-Congo component)
KEN_Kakapel_300BP (ancient Nilo-Saharan component)
MWI_Chencherere (ancient East African Hunter-Gatherer component)
ZAF_2000BP (ancient Khoisan component)
Least-admixed Iberomaurusian reference sample:
MAR_Taforalt (Iberomaurusian component)
Least-admixed East Eurasian reference sample:
CHN_Huatuyan_500BP (ancient East Asian component)
Least-admixed South Eurasian reference sample:
LAO_Hoabinhian (Ancient Ancestral South Indian component)
The resulting analytical table is as follows:





























From the data tables above, we may note that:








In summation, the genomic evidence indicates that the present-day Semitic-speaking populations of the Middle East may be divided into seven basic groups based on their ancestral composition:
One data point which stands out is the very heterogeneous nature of the modern Syrian population. Moreover, the fact that all but two of the Semitic samples lacked the MWI_Chencherere component of the ancient East African Hunter-Gatherers is quite telling. Since this population element is found at an average frequency of 8.6% among Afro-Asiatic-speaking populations of the Horn (see here), this reveals that the Cushitic component that certain researchers have detected at low frequency in the genomes of some contemporary Middle Eastern Semitic speakers (<10%) was specifically derived from Sudanese “Arabs” and/or Nubians (i.e. Cushitic-related peoples from the Nile Valley, where East African Hunter-Gatherer admixture is practically non-existent and Nilo-Saharan admixture is instead more pronounced; cf. Gopalan et al. (2019)). Finally, the presence of the Iberomaurusian/Taforalt component at highest frequencies among the Ashkenazi Jew samples suggests a connection with North Africa. Such a link is also supported by these individuals’ excess Anatolian Neolithic ancestry, which has long been a common ancestral element in the Maghreb.
Genotype and phenotype of the Middle-to-Late Bronze Age Semites of Alalakh
How and when the Ashkenazi Jew and Sephardic Jew gene pool was formed
Brief note on the Levantine origin of Ashkenazi and Sephardic Jews
The difference between “pure” Semites (Mahra) and Afro-Arabs (Akhdam)
Y-DNA haplogroup E1b1a (formerly E3a) appears to have originated in the Middle East
Guide to the Universal Basic Genetic Model
23andme and AncestryDNA misrepresent the ancestral composition of Afro-Asiatic speakers
Phenotype of the Canaanites, ancestors of modern Semitic peoples of the Levant and Mesopotamia
Genetic and phenotypic affinities of the Bronze Age Semites of Sidon, Lebanon
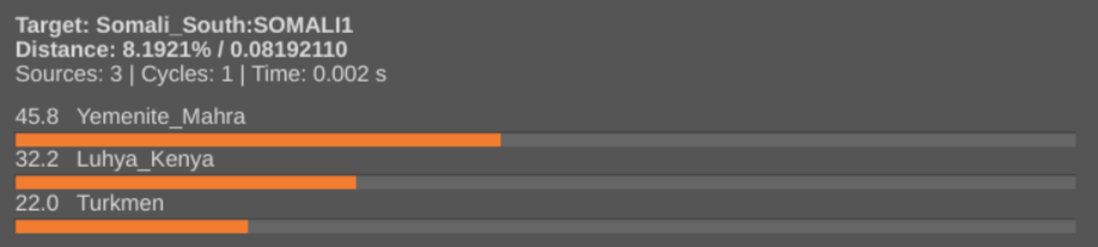
On Detecting the Eurasiatic genome component in Cushitic peoples and European Steppe-associated mtDNA haplogroups among the Cushitic peoples, I explained how, in addition to their Levantine Natufian ancestry, Cushitic individuals carry European Steppe and ancient East Asian ancestral elements. I have termed these latter components “Eurasiatic” because among Afro-Asiatic speakers in Northeast Africa, these elements always appear together and at roughly the same ratio, as though they had been spread by/inherited from an ancient population of mixed Steppe-East Asian extraction (e.g. the Turkmen of Central Asia — see Appendix below).
A reader has courteously pointed out to me that there is also apparently some linguistic evidence, which further supports an early Eurasiatic presence in the Nile Valley and Horn of Africa.
The scholar Wilfred H. Schoff (1912) notes that the Cushitic languages of Northeast Africa are related to the ancient Egyptian and Semitic tongues (we now know that this is because these idioms all belong to the Afro-Asiatic family). Controversially, he also claims that, in terms of syntax, Oromo, Somali, Beja and other Cushitic languages of the Horn and Nile Valley share some commonalities with the Altaic tongues of Central Asia, such as the Turkic language spoken by the aforementioned Turkmen:
[…]in the so-called Cushite languages of northeast Africa, such as the Galla, Somali, Beja, and other allied dialects, we find grammatical principles analogous to those of the early Egyptian and Semitic tongues combined with a totally dissimilar syntax presenting no analogy with that of the Semites or with any Negro tongue in Africa, but resembling closely the syntax of the Ural-altaic languages of Asia[…]
This is the first time I am hearing of a possible Altaic influence on the Cushitic languages. If true, it wouldn’t really surprise me given the fact that Cushitic peoples do bear European Steppe and ancient East Asian genome components, like those which typify many Turkic speakers in Central Asia.
Genome analysis of Mesolithic to Bronze Age individuals in the Levant, moreover, indicates a near complete absence of European Steppe and ancient East Asian ancestries. Ergo, whichever ancient population(s) spread these Eurasiatic ancestral elements in Northeast Africa likely came from elsewhere and did not speak an Afro-Asiatic language(s). These outsiders very well could instead have spoken an Altaic idiom or even an Indo-European tongue.




Detecting the Eurasiatic genome component in Cushitic peoples
European Steppe-associated mtDNA haplogroups among the Cushitic peoples
The biological affinities of the Cushitic peoples, both genetic and physical, is a topic that seems to attract a lot of interest.
On Detecting the Eurasiatic genome component in Cushitic peoples and Correcting a faulty genetic model #1, we have seen how, when properly genetically modelled, Cushitic peoples predominantly bear non-African ancestry (over 70% on average). This non-African heritage consists of a Levantine Natufian genome component and Eurasiatic genome components (that is, European Steppe and ancient East Asian ancestral elements). Additionally, these individuals carry minor Sub-Saharan African admixture (~25%), and trace North African Iberomaurusian admixture (under 5%).
Furthermore, we know that in their “pure” (i.e. unadmixed) form, modern Cushitic peoples generally have a “Caucasoid” craniofacial pattern and soft-textured hair. Some Cushitic individuals also have a very light complexion, referred to locally as cad. Moreover, a significant minority of contemporary Cushitic individuals possess light-colored eyes and hair.
But where do these traits actually come from? Can most or all of them be traced back to the ancient Cushites themselves? Or are they instead a product of recent admixture with outsiders (e.g. Persians, Europeans, etc.?).
Let us examine this issue in detail using scientific data.
Wang et al. (2020) examined two of the ancient Cushitic settlers of the Pastoral Neolithic for phenotype alleles; a 2300 year old sample from Hyrax Hill and a 1500 year old sample from Molo Cave, archaeological sites located in Kenya. The scientists report that both of the specimens bore the derived Ala111Thr (rs1426654) allele of the SLC24A5 gene, which confers a lighter complexion (cf. Table S7). Of the known genetic variants controlling skin pigmentation (which number over 20), this derived SLC24A5 mutation has been measured to have the single greatest effect on coloration: 7.6-11.4 average melanin units for the derived SLC24A5 allele, compared to just 5 average melanin units for the derived SLC45A2 allele and 2.8-3.8 average melanin units for the derived KITLG allele (Beleza et al. (2013)).
Besides the derived SLC24A5 variant, Wang et al. observed that their ancient Cushitic samples carried the derived allele at the APBA2 locus, which is likewise associated with lighter skin pigmentation.
Altogether, ancient DNA analysis suggests that the early Cushites would have been of a relatively fair complexion, though just how light-skinned is uncertain since these individuals were not analysed for the other, less impactful pigmentation-related alleles. This, in turn, means that the ancient Cushites did not inherit their lighter skin-conferring mutations from the Levantine Natufians because the Natufians did not have any such alleles.
Genetic analysis of modern Cushitic, Ethiosemitic and North Omotic speakers of the Horn indicates that most carry the derived SLC24A5 allele (~60%; cf. ALFRED; et al. (2015)). These Afro-Asiatic-speaking individuals have not been studied for the derived APBA2 mutation of the early Cushites. However, they have been found to bear low frequencies of the derived SLC45A2 allele (0%-1.4% of Ethiopian Jews; 0%-3.3% of Somalis), as do peninsular Arabs (10.5% of Qataris; 11.6% of Saudis) (cf. ALFRED).
This is similar to ancient Egyptian specimens excavated at the Abusir site in Middle Egypt, two of whom Schuenemann et al. (2017) assert carried the derived allele for the SLC24A5 gene but not the derived SLC45A2 mutation. The derived SLC45A2 allele seems instead to have been mainly spread in the Levant and Arabian peninsula during the later Bronze Age by foreigners from the Caucasus/Iranian Plateau since: a) Bronze Age specimens exhumed at Sidon, Lebanon, who are otherwise very similar to modern Lebanese individuals, did not bear this variant (Haber et al. (2017)), and b) this mutation is today found at elevated frequencies among Levantine populations that have appreciable Caucasus Hunter-Gatherer-related admixture (Druze, Samaritans).

Examination of the bones of ancient Cushitic settlers in eastern Africa has established that, on average, they had a slender constitution. This was a distinctive characteristic they shared with their relatives the ancient Egyptians.
In his capacity as chief anatomist for the Archaeological Survey of Nubia, the Egyptologist Grafton Elliot Smith inspected thousands of predynastic Egyptian specimens. He also studied many predynastic Egyptian skeletons excavated from the Naqada site by the Hearst Expedition under George Andrew Reisner, as well as predynastic Egyptian remains recovered by Randall-MacIver at El-Amrah near Abydos. According to Elliot Smith (1911):
The hot, dry sands of Egypt have preserved through a span of more than sixty centuries the remains of countless multitudes of the earliest people known to have dwelt in the Nile Valley ; and not the mere bones only, but also the skin and hair, the muscles and organs of the body ; and even such delicate tissues as the nerves and brain, and, most marvellous of all, the lens of the eye, are available for examination to-day. Thus we are able to form a very precise idea of the structure of the body of the Proto-Egyptian.[…]
Our information concerning these earliest inhabitants of the Nile Valley has been acquired from the study of the contents of many thousands of their graves, found in cemeteries scattered in every part of Egypt and Nubia so far examined.[…]
[The Proto-Egyptian] was of very slender build, for his bones are singularly slight and free from pronounced roughness and projecting bosses that indicate great muscular development. In fact, there is a suggestion of effeminate grace and frailty about his bones, which is lacking in the more rugged outlines of the skeletons of his more virile successors.
Thus, modern Cushitic, Ethiosemitic and North Omotic-speaking individuals of Northeast Africa simply inherited their linearity from their Cushitic ancestors.
We know with some certainty that this physique was not acquired from ancient Nilo-Saharan peoples. For starters, anthropometric analyses have shown that contemporary Afro-Asiatic speakers from the Horn (viz. Somalis, Oromos/Gallas, Afars, Amhara) cluster with Egyptians, Nubians and Yemenis rather than with Nilo-Saharan speakers from the Nile Valley (Mabaan, Nuba, Dinka, Shilluk, Anuak) (cf. (Billy (1988); Leguebe (1981)).


Furthermore, Melton et al. (2002) found that Cushitic Somalis have lumbar spine and femoral neck bone mineral content, bone mineral density, and bone mineral apparent density that is overall more similar to that of European Americans than to that of African Americans. By contrast, individuals from south Sudan (most of whom are Nilotes) have lumbar spine bone mineral density that is significantly lower than that of both European Americans and African Americans (Gong et al. (2006)).

Forensic examination of the ancient peoples of the Kerma civilization in Sudan indicates that, except for a few assimilated individuals of Nilotic origin, they uniformly had non-kinky hair texture. Lepisus (1915) notes that “most of the men, especially the principal skeletons, had fine heads with straight black hair.” According to linguistic analysis by Marianne Bechaus-Gerst (1989, 2000), these Kerma folks spoke languages from the Cushitic branch of the Afro-Asiatic family.

Furthermore, Wang et al. (2022) genetically analysed one such Kerma period specimen, who was excavated at the Kadruka site in Upper Nubia (northern Sudan). This individual had soft-textured wavy hair. His genome was found to be “genetically indistinguishable from that of early Neolithic eastern African pastoralists located 2500 kms away.” Ergo, the Cushites of the Pastoral Neolithic, who bore a predominant non-African ancestry (see above), passed on their soft-textured hair to their modern descendants.

As I wrote on Origin and incidence of light-colored eyes among the Cushitic peoples:
It is not known whether or not the Pastoral Neolithic Cushites specifically had blue, grey, green or hazel eyes. However, based on genetic analysis of ancient Cushitic specimens, what is known is that some of them probably did have light-colored eyes.
Wang et al. (2020) report that two of the Pastoral Neolithic Cushitic individuals they examined, who were excavated from the Lukenya Hill site in Kenya, both carried the derived rs1800404 allele at the OCA2/HERC2 locus. This variant is associated with lighter eye pigmentation (cf. Table S7).
Having said that, it would appear that another SNP, the rs916977 mutation of the HERC2 gene, is one of the main alleles responsible for the instances of light eye coloration found among modern Cushitic, Ethiosemitic and North Omotic speakers. This variant is borne today by around 27% of Ethiopian Jews (cf. ALFRED). Interestingly, almost an equal percentage of Sandawe individuals in eastern Africa have been found to also carry this mutation. Since the Sandawe are known to have assimilated many Cushitic pastoralists, this suggests that the earlier Cushitic peoples they interacted with (and from whom they presumably obtained the rs916977 allele) bore the SNP at elevated frequencies.

Archaeogenetic analysis indicates that the Cushites of the Pastoral Neolithic were relatively fair-skinned. These individuals bore derived variants at the SLC24A5 and APBA2 loci, which are known to confer lighter pigmentation. Physical examination of the Cushites’ relatives the ancient Egyptians and modern Cushitic individuals further suggests that they had a slender build. Additionally, forensic examination of early Cushitic specimens has generally found that they had soft-textured hair. Although the Cushites of the Pastoral Neolithic were not analysed for alleles related to hair pigmentation, two Cushitic individuals that were examined at the Lukenya Hill site were both found to carry the derived rs1800404 allele of the OCA2/HERC2 gene, a mutation which is associated with light eye coloration. These results are consistent with the genome affinities of ancient and modern Cushitic individuals. In particular, the fact that Cushitic peoples from a period as early as the Neolithic already bore specific variants associated with light skin (APBA2) and light eyes (rs1800404) supports the idea that these alleles were inherited from the European Steppe side of Cushitic ancestry. These mutations were clearly not inherited from the Levantine Natufians since the Natufians did not carry any alleles associated with lighter skin, hair or eye pigmentation. They also do not seem to have been inherited from the Natufians’ immediate descendants, the Pre-Pottery Neolithic (PPN) makers, since the PPN folks bore elevated Anatolian Neolithic admixture, an element which the coeval Cushites of the Pastoral Neolithic did not possess.
Detecting the Eurasiatic genome component in Cushitic peoples
Correcting a faulty genetic model #1
Correcting a faulty genetic model #2
Northern Somalis from Puntland (Majerteen Darod) have the most non-African ancestry in the Horn
Hair form among ancient and modern Cushitic peoples
Origin and incidence of light-colored eyes among the Cushitic peoples
Cushites in the northern Horn are “purer” than Cushites in the southern Horn
A note on the mariin and cad phenotypes, the original physiognomies of the Cushitic peoples
Guide to the Universal Basic Genetic Model
Phenotype of the Canaanites, ancestors of modern Semitic peoples of the Levant and Mesopotamia
Genetic and phenotypic affinities of the Bronze Age Semites of Sidon, Lebanon
Ancient Khoisan were dark-skinned and did not look Eurasiatic
On Phenotype of the Canaanites, ancestors of modern Semitic peoples of the Levant and Mesopotamia, we saw how the Bronze Age Canaanites of Israel/Palestine, were dark-skinned, dark-eyed and dark-haired. No Canaanite individuals were pale-skinned, blue-eyed or red-haired/blonde-haired. This result is consistent with the ultimate Levantine Natufian ancestral roots of the Semitic peoples, as the Natufians did not carry alleles conferring light skin, light eye or light hair pigmentation.
But what about the Canaanite-related inhabitants of Sidon, Lebanon, another Semitic population of the Bronze Age? Were they also swarthy?
Let us take a closer look and find out.
Haber et al. (2017) genetically examined ancient Sidon individuals. Performing a Principal Component Analysis, they observed that the Sidon samples clustered with present-day Levantines. This suggests that the Bronze Age inhabitants of Sidon were immediately ancestral to modern Lebanese and other Levantine individuals:
The Bronze Age Sidon samples (Sidon_BA) overlap with present-day Levantines and were positioned between the ancient Levantines (Natufians/Neolithic) and ancient Iranians (Neolithic/Chalcolithic). The overlap between the Bronze Age and present-day Levantines suggests a degree of genetic continuity in the region.

Haber et al. also surveyed the Sidon samples for phenotypic alleles. They observed that these ancient Semitic individuals bore the SLC24A5 variant associated with lighter skin pigmentation. This mutation appears to have been introduced into the local Semitic gene pool by foreigners from the Iranian plateau, who bore Iran Neolithic-associated ancestry and the Y-DNA haplogroup J:
In addition, the two Sidon_BA males carried the Y-chromosome haplogroups45 J-P58 (J1a2b) and J-M12 (J2b) (Tables 1 and S4; Figure S11), both common male lineages in the Near East today. Haplogroup J-P58 is frequent in the Arabian peninsula with proposed origins in the Zagros/Taurus mountain region. It forms the vast majority of the Y chromosomes in southwestern Mesopotamia and reaches particularly high frequencies (74.1%) in Marsh Arabs in Iraq. On the other hand, haplogroup J-M12 is widespread at low frequency from the Balkans to India and the Himalayas, with Albanians having the highest proportions (14.3%). We compiled frequencies of Y-chromosome haplogroups in this geographical area and their changes over time in a dataset of ancient and modern Levantine populations (Figure S12), and note, similarly to Lazaridis et al., that haplogroup J was absent in all Natufian and Neolithic Levant male individuals examined thus far, but emerged during the Bronze Age in Lebanon and Jordan along with ancestry related to Iran_ChL. All five Sidon_BA individuals had different mitochondrial DNA haplotypes (Table 1), belonging to paragroups common in present-day Lebanon and nearby regions (Table S5) but with additional derived variants not observed in our present-day Lebanese dataset.
However, unlike many peoples today in the Levant, the ancient Sidon individuals did not also bear the SLC45A2 variant nor apparently any other light skin-associated mutations.
Altogether, this suggests that the occupants of Sidon were more deeply pigmented than their modern descendants. It also indicates that, since the Bronze Age, Levantine individuals acquired additional light skin-associated variants through contact with outsiders:
SNPs associated with phenotypic traits show that Sidon_BA and the Lebanese had comparable skin, hair, and eye colors[…] but with Sidon_BA probably having darker skin than Lebanese today from variants in SLC45A2 resulting in darker pigmentation.

Genome analysis of modern Semitic individuals
Phenotype of the Canaanites, ancestors of modern Semitic peoples of the Levant and Mesopotamia
Genotype and phenotype of the Middle-to-Late Bronze Age Semites of Alalakh
For a change, we shall explore today some widely-held myths about the Afro-Asiatic-speaking populations in light of modern science.
Genome analysis of ancient Egyptian mummies dating from the Old Kingdom indicates that they mostly bore Levantine Natufian ancestry. Later dynastic Egyptians from the Old Kingdom-Middle Kingdom carried less Levantine Natufian ancestry, primarily due to admixture with outsiders bearing Caucasus Hunter-Gatherer ancestry. Modern Egyptians (Coptic and Muslim alike) directly descend from the ancient Egyptians of the Old Kingdom-Middle Kingdom period.
For details, see The Dynastic Race Theory might be true.



Genome analysis indicates that Cushitic, Ethiosemitic and North Omotic speakers of the Horn of Africa predominantly carry non-African ancestry (over 70% on average). This non-African heritage consists of a Levantine Natufian component and European Steppe and ancient East Asian components. They also have minor Sub-Saharan African admixture (~25%), and trace North African Iberomaurusian admixture (under 5%).
Bad genetic modelling is the main reason why some researchers mistakenly believe that Cushitic peoples are a product of Arabians mating with Sub-Saharan Africans. These faulty genetic models have one or more of the following three errors in common:
For details, see Detecting the Eurasiatic genome component in Cushitic peoples, Correcting a faulty genetic model #1 and Correcting a faulty genetic model #2.



Genetic studies indicate that a gradient of non-African ancestry exists among the Afro-Asiatic-speaking groups in the Horn vicinity. Principal Component Analysis suggests that, on average, this non-African ancestry peaks among Northern Somali individuals, who hail from the northeastern Puntland region in Somalia (mainly Majerteen Darod). Proximal genome analyses further reveal that many Cushitic-speaking individuals in southern Somalia, southern Ethiopia and the Great Lakes region have an additional pulse of Sub-Saharan African admixture, which is generally not borne by modern Cushitic peoples in the northern Horn nor the ancient Cushites of the Pastoral Neolithic.
For details, see Northern Somalis from Puntland (Majerteen Darod) have the most non-African ancestry in the Horn.


The Amharic and Tigrinya Ethiosemitic languages both contain a strong Agaw substratum, retained from the Central Cushitic languages which the early Abyssinians spoke. Culturally, Agaws and Abyssinians (Habesha) are also very similar.
Furthermore, genome analysis indicates that Cushitic-speaking Agaws and Ethiosemitic-speaking Abyssinians share a Cushitic ancestral base. Both groups of individuals not only carry the same overall ancestral components and at similar percentages, they also derive most of their Levantine Natufian ancestry from a direct Natufian population source.
For details, see Genetic analysis of the Cushitic-speaking Agaws (Agaw proper) and Semitic-speaking Agaws (Abyssinians/Habesha).




Generally-speaking, Sudanese “Arabs” are of the same ancestral origin as Nubians. They share close genetic, physical and (to some extent) cultural ties. Textual evidence also indicates that they spoke a common language as recently as the Middle Ages, during the spread of Islam. This is when Kababish, Gaalien, Arakien, Meseria and other Sudanese “Arab” groups are believed to have adopted their current Arabic language and Arabian genealogies. Consequently, in genetic distance analyses which investigate the proximal or recent ancestry of individuals, Sudanese “Arabs” show a clear preference for the medieval Kulubnarti Nubian specimens as opposed to recent peninsular Arab specimens (such as Levant_Tell_Qarassa_Early_Antiquity).
Overall, the northern Sudanese — that is, Nubians and Sudanese “Arabs” — have a similar ancestral composition as the Afro-Asiatic-speaking populations from the Horn. The Kababish “Arabs” of Sudan carry a predominant non-African ancestry (averaging almost 70%), which consists of Levantine Natufian, European Steppe and ancient East Asian components. They also bear some Sub-Saharan African admixture (nearly 30% on average), which varies considerably per individual, much more than among the Afro-Asiatic speakers of the Horn. Certain Kababish individuals have a low SSA admixture of ~20%, whereas others have a high SSA admixture of ~50%.
For details, see First genomes from the ancient Sahara detect two different groups of pastoralist inhabitants.



Most Arabic speakers in the Maghreb are Arabized Berbers. This has been confirmed genetically, as they carry the same overall signature Maghrebi components as Berber speakers. These ancestries mostly consist of North African Iberomaurusian, Anatolian Neolithic and Levantine Natufian elements. Additionally, these individuals have minor Sub-Saharan African admixture.
The recent peninsular Arab ancestry found among Northwest Africans (best represented by the Levant_Tell_Qarassa_Early_Antiquity component) is generally low, comprising an average of ~10% of their ancestral makeup.
Having said that, there are some groups and individuals in Northwest Africa that clearly are descended from recent settlers from the Arabian peninsula. These communities include the Rbaya of Tunisia. Many individuals inhabiting the Douz area, also in Tunisia, likewise seem to be of recent peninsular Arab origin.
For details, see Guide to the Universal Basic Genetic Model.



Genome analysis indicates that among all modern populations, the Yemenite Mahra, Socotri and other southern Arabians carry the most Levantine Natufian ancestry. Contrary to popular belief, these individuals bear little-to-no Sub-Saharan African or Ancient Ancestral South Indian admixtures. They are, quite literally, the “purest” (i.e. the least admixed) living Semitic peoples.
For details, see The difference between “pure” Semites (Mahra) and Afro-Arabs (Akhdam) and Medieval DNA from Socotra confirms that the Socotri are among the “purest” descendants of the Levantine Natufians.




Genetic analysis indicates that Ashkenazi Jews and Sephardic Jews descend from Middle Eastern Jews, who settled in southern Europe. There, their Levantine ancestors (primarily men) intermixed with autochthonous Europeans (primarily women), and subsequently dispersed to other parts of the continent. This is why Ashkenazi and Sephardic Jews today have mostly Levantine-related Y-DNA or paternal DNA (derived from their Semitic male ancestors) and mostly southern European-related mtDNA or maternal DNA (derived from absorbed indigenous European women).
In terms of genome affinities or autosomal DNA, this is also why modern Ashkenazi and Sephardic Jewish individuals are intermediate between Levantine and southern European populations. On average, they bear around 50% Lebanese-related ancestry and 50% north Italian-related ancestry, like contemporary Maltese persons and Sicilians.
For details, see Ashkenazi Jews and Sephardic Jews are of the same Levantine ancestral origin as Mizrahi Jews, Palestinians and other Semites Brief note on the Levantine origin of Ashkenazi and Sephardic Jews, and Genome analysis of modern Semitic individuals.


The Dynastic Race Theory might be true
Detecting the Eurasiatic genome component in Cushitic peoples
Correcting a faulty genetic model #1
Correcting a faulty genetic model #2
Northern Somalis from Puntland (Majerteen Darod) have the most non-African ancestry in the Horn
First genomes from the ancient Sahara detect two different groups of pastoralist inhabitants
Guide to the Universal Basic Genetic Model
The difference between “pure” Semites (Mahra) and Afro-Arabs (Akhdam)
Brief note on the Levantine origin of Ashkenazi and Sephardic Jews
Phenotype of the Canaanites, ancestors of modern Semitic peoples of the Levant and Mesopotamia
Genetic and phenotypic affinities of the Bronze Age Semites of Sidon, Lebanon
Canaanites were a Bronze Age people of the Levant, descended from the Pre-Pottery Neolithic (PPN) culture bearers. The PPN makers, in turn, immediately descend from the Natufians of the Mesolithic period. Ergo, the Pre-Pottery Neolithic folks serve as a link between the Canaanites and their Natufian ancestors. The Canaanites and other coeval populations of the Levant and Mesopotamia (e.g. the inhabitants of Alalakh, Ebla and Sidon) likewise connect modern Semitic peoples with their Pre-Pottery Neolithic forebears and, more remotely, with their Natufian progenitors.
Because a) the Canaanites hold a key position within Semitic ethnogenesis, and b) the ancient land of Canaan was located in the disputed territory of present-day Israel/Palestine, the biological affinities of the Canaanites have been hotly debated.
Let us examine one key aspect of that controversy, the phenotypic affinities of the ancient Canaanites.



Lazaridis et al. (2022) conducted a comprehensive analysis of phenotypic traits borne by ancient individuals excavated in Europe and Asia, including the Middle-to-Late Bronze Age peoples of Canaan.
With regard to skin color, the scientists report that most of the Canaanites were deeply pigmented (71%), having either a dark skin tone (4/7 specimens or 57%) or a dark-to-black skin tone (1/7 specimens or 14%). A minority of Canaanite individuals had an intermediate skin tone (2/7 specimens or 29%). None of the Canaanite individuals reportedly had a pale skin tone (0/7 specimens or 0%).
Similarly, in terms of hair color, Lazaridis et al. note that all of the Canaanites had darker manes (7/7 or 100%). Of these individuals, most had black hair (5/7 specimens or 71%) and a few had brown hair (2/7 or 29%). None of the Canaanite individuals had either red hair or blond hair.
Finally, with respect to eye color, all of the Canaanite individuals had brown eyes (7/7 or 100%). None were reported to possess blue eyes.


The phenotypic affinities of the Canaanites will hardly come as a surprise to readers of this website. As I have repeatedly pointed out, Natufian and Iberomaurusian ancestries are associated with darker skin pigmentation because the ancient Natufians and their Iberomaurusian relatives did not carry alleles which confer lighter skin. This is why the modern individuals with the most Natufian ancestry (viz., Yemenite Mahra, Socotri, etc.) are all dark brown-to-black in color, despite possessing around 20% of Caucasus Hunter-Gatherer admixture (an ancestral element which does, by contrast, contain alleles conferring lighter skin pigmentation).
The dark-skinned, dark-haired and dark-eyed appearance of the Natufian-descended Canaanites, now confirmed by empirical genetic data, explains several things at once:



Genome analysis of modern Semitic individuals
Genotype and phenotype of the Middle-to-Late Bronze Age Semites of Alalakh
Brief note on the Levantine origin of Ashkenazi and Sephardic Jews
Guide to the Universal Basic Genetic Model
Genetic and phenotypic affinities of the Bronze Age Semites of Sidon, Lebanon
I have read some propaganda online claiming that the Tutsi, a Bantu-speaking people living in Central-East Africa, are of Cushitic origin. This could not be further from the truth. The Tutsi are actually of Nilotic origin, as should be obvious from their “Negroid” appearance and close physical resemblance to the Maasai and other Cushitic-admixed Nilotes inhabiting the Great Lakes region.

Craniometrically, the anthropologist G. P. Rightmire (1975) observed that his Iron Age Rwanda male and female samples cluster with his Bantu and Khoisan samples. All of these specimens are also morphologically distinct from the ancient Cushitic male and female samples, which instead cluster with the ancient Egyptian male and female samples (Baharini and Makalia in particular):

Furthermore, because the Tutsis’ Bantu ancestors in the past assimilated some Southern Cushites, the average Tutsi individual today tends to have narrower facial features than other Niger-Congo/Nilo-Saharan speakers. However, the nasal width of the Tutsi (estimated by Jean Hiernaux (1974) at 39 mm, similar to the Maasai Nilotes) remains significantly broader than that of Afro-Asiatic speakers (estimated at 36 mm for the Warsangali Darod Somali of northern Somalia).
Likewise, David Blackwell (1984) photogrammatically analysed the nasal morphology of the Tutsi Bantus as compared to that of the Hehe Bantus and found that these Niger-Congo-speaking populations are similar to each other on most indices, including nasal prominence, nasal bridge distance, columella length, nasal height, nasal angle, nasal wing/septum relationship, and nasal region vertical relationship. Moreover, although the Tutsis generally have a lower nasal index than their Hehe brethren, the Tutsi also have a considerably larger nasal breadth index and inter-occular nasal width index. This “again suggests the Tutsi’s noses are wider relative to face width than the Hehe’s”.
Additionally, Hiernaux (1974) reports that all Tutsis have kinky hair texture like the Hutu Bantus, Maasai Nilotes and other Niger-Congo, Nilo-Saharan and Khoisan populations.
Altogether, this tells us that the Tutsi’s Iron Age ancestors were not Cushitic people, but instead morphologically “Negroid” and thus similar in physiognomy to other Bantus, Nilotes and Khoisan.
Serological (blood group) analyses indicate that, overall, the Tutsis are similar to other Bantus/Nilotes. Lawrence Oschinsky (1960) states:
The Batutsi are tall, narrow nosed, narrow faced Negroids showing evidence of previous Caucasoid admixture, the Bahutu are intermediate in stature and have broader noses and faces, and the Batwa represent a local variant of Congo pygmy racial type. In the ABO, Rh and MN blood group distributions the Batutsi and the Bahutu are similar.
J. D. Fage (2013) likewise notes that:
The origin of the Tutsi/Hima/Chwezi ruling class in the lacustrine Bantu kingdoms is an intriguing question. Serologically they are Blacks, and this seems to rule out the possibility of a Cushitic origin.
Genetically, the Tutsis again show close ties with the Maasai Nilotes and Hutu Bantus.
Peer-reviewed studies report that 75% of Tutsi individuals in Rwanda and Burundi carry the E1b1a haplogroup, a clade formerly known as E3a (Trombetta et al. (2015); Luis et al. (2004); Luis et al. (2007)). This is the most common paternal lineage borne today by the Hutu Bantus and other Niger-Congo-speaking individuals, as well as most Maasai. Only 1% of Tutsis in Rwanda and 22% of Tutsis in Burundi have been found to harbor M296, a subclade of the Afro-Asiatic-affiliated haplogroup E1b1b (formerly E3b), which is associated with the ancient spread of Southern Cushites:


In terms of mtDNA, Göbel et al. (2019) report that almost all of their Rwanda sample belongs to derivatives of the macrohaplogroup L. Much of this mitochondrial variation was also shared with the Kenya cohort. The scientists did, however, detect a minor Cushitic influence in their Rwandan dataset via the presence of the West Eurasian maternal lineages M1a1 (2.6%), N1a1a (1.3%) and K1a (0.6%) as well as the U6a clade.
For their overall ancestry, all studies that have examined the autosomal DNA of Tutsi individuals indicate that their STR markers are most similar to those of the Hutu and other Niger-Congo-speaking populations (cf. Coudray et al. (2006); Simms et al. (2008); Shepard and Herrera (2006b); Shepard and Herrera (2006a)):

Miao et al. (2024) compared the genomes of Rwandan individuals with those of various global populations. They observed that their Rwandan sample clustered with their Nilote Luo sample from Kenya, consistent with the Nilotic roots of the Tutsi. According to the researchers:
Rwanda is known as the heart of Africa, reflecting the history of the world. Colonization and genocide have led to Rwanda’s existing genetic structure. Herein, we used massively parallel sequencing to analyze 296 loci in 185 Rwandans and constructed a database for Rwandan forensic data for the first time. We found the following results: First, forensic parameters demonstrated that all loci were highly informative and could be used for forensic identification and paternity tests in Rwandans. Second, we found that the differences in genetic background between Rwandans and other African populations were similar but slight, as indicated by the massively parallel sequencing panel. Rwandans belonged to the African population and were inseparable from populations from neighboring countries. Also, Rwandans were closer to the European and American populations because of colonization, war, and other reasons. There was no scientific basis for racial classification established by colonization. Further research still needs to be carried out on more loci and larger Rwandan samples.

Autosomal SNP analysis by 23andme, AncestryDNA and other commercial genome testing labs similarly report that Tutsi individuals share most of their ancestry (close to 70%) with Niger-Congo/Nilo-Saharan/Khoisan individuals. Of this predominantly Sub-Saharan African ancestry, much of it is associated with Southern East African Bantus (44.8% for the Rwandan Tutsi individual below, tested with 23andme). They also have some admixture from Nilotic peoples from the Sudan area (16.2% below). Additionally, Tutsis bear admixture derived from Cushitic peoples of Ethiopia and Eritrea (34.2% below), whom their Nilotic/Bantu ancestors absorbed:
 Finally, genome analysis of Tutsi individuals using the Vahaduo Admixture JS program indicates that, like the Maasai and other Nilotic peoples inhabiting the Great Lakes region, they trace most of their proximal or recent ancestry to the Cushitic-admixed Nilotes of the Pastoral Iron Age. This confirms that the Tutsi are indeed of Nilotic origin since all peoples of genuine Cushitic origin (such as the Somali, Oromo, Afar, Agaw/Abyssinians) trace instead their proximal ancestry to either the Cushites of the Pastoral Neolithic or the Cushitic-related inhabitants of medieval Kulubnarti. Genetic distance analysis also indicates that the Tutsi and Hima share closest genetic affinity with the Nilotic Ogiek people of Kenya and other mostly Nilotic/Bantu populations, further confirming the Nilotic origins of the Tutsi-Hima.
Finally, genome analysis of Tutsi individuals using the Vahaduo Admixture JS program indicates that, like the Maasai and other Nilotic peoples inhabiting the Great Lakes region, they trace most of their proximal or recent ancestry to the Cushitic-admixed Nilotes of the Pastoral Iron Age. This confirms that the Tutsi are indeed of Nilotic origin since all peoples of genuine Cushitic origin (such as the Somali, Oromo, Afar, Agaw/Abyssinians) trace instead their proximal ancestry to either the Cushites of the Pastoral Neolithic or the Cushitic-related inhabitants of medieval Kulubnarti. Genetic distance analysis also indicates that the Tutsi and Hima share closest genetic affinity with the Nilotic Ogiek people of Kenya and other mostly Nilotic/Bantu populations, further confirming the Nilotic origins of the Tutsi-Hima.



For a complete biological analysis of the Tutsi Bantus, I recommend the excellent essay The Elongated African fallacy on the Land of Punt website. It covers all of the above and much more in detail, including the Tutsi’s HLA antigen affinities and other classical genetic markers.
Tutsi men and women:





























































Among affluent Tutsi individuals and models, skin bleaching/lightening and the wearing of hair weaves or chemical hair straightening is relatively common. These cosmetic alterations serve to obscure the Tutsis’ predominant Nilotic/Bantu ancestry (over 70%), while highlighting their minority Cushitic admixture (under 30%). This practice can be seen below in the person of Ange Kagame, a Tutsi individual, and daughter of the Rwandan president Paul Kagame:


Overall, the Tutsi closely resemble the Hutu, another Bantu-speaking population inhabiting Rwanda and Burundi.
Tutsi and Hutu men, women and children:









Maasai Nilotic men and women — note the close physical resemblance to the Tutsi Bantus, stemming from their shared Nilo-Saharan origins:




A very Eurasiatic Toubou sample
Baggara/Shuwa “Arabs”: last remaining descendants of the original Chadic speakers
Anthropometric analysis of the Cushitic-speaking Beja and Iraqw

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}